Генетика тикированных окрасов кошек.
Прежде чем рассматривать проблему генетического взаимодействия тикинга и типпинга, определимся в терминах. Тикинг (ticking)- зонарное окрашивание волоса, которое образуется при чередовании двух откладывающихся пигментов – черного и желтого. Проявляется этот признак только при наличии в генотипе животного агути-фактора, то есть доминантного аллеля гена Agouti. Тикированными тэбби называются кошки, у которых остевой и покровный волос на корпусе имеет равномерное зонарное окрашивание, благодаря чему рисунок на теле животного отсутствует. Самый яркий пример тикированного окраса – абиссинские кошки, у которых нет рисунка не только на корпусе, но также и на ногах и хвосте. Типпинг (tipping) – окрашивание верхней части волоса при полной или частичной депигментации его нижней части. При полном отсутствии пигмента в нижней части волоса эта последняя выглядит белой (т.наз. “серебро”), при отсутствии в ней только черного пигмента – желтой или абрикосовой (“золото”). Традиционно подразумевается, что верхняя часть типпированного волоса окрашена в черный цвет: если большая часть остевой и покровной шерсти прокрашена на 1/3, окрас называется затушеванным, если на 1/8 - затененным или “шиншилла”. Любой из этих окрасов, между которыми, кстати, нет четкой границы, вполне корректно будет назвать типпированным. Для проявления типпированных окрасов так же, как и для развития тикинга, необходимо наличие у животного агути-фактора.
Долгое время тикинг и типпинг трактовались как взаимоисключающие явления. Однако такое положение верно только для стандартов окраса. В реальности же наличие у кошки типпинга отнюдь не исключает одновременного присутствия тикинга: верхняя часть волоса может быть зонарно окрашена, а нижняя – депигментирована. Правда, существование таких животных долгое время “выпадало” из поля зрения заводчиков. Поскольку их окрас не был описан в стандарте и являлся лишь нежелательным отклонением от оного, заводчики предпочитали этих кошек… не замечать.
Генетика типпинга: исторический обзор
Первыми типпированными кошками, “обнаруженными” фелинологами, были серебристые шиншиллы и затушеванные. Соответственно, необходимым для формирования типпированных окрасов был признан (помимо уже упоминавшегося агути-фактора) ген – ингибитор меланина. Действие доминантного аллеля этого гена ясно из самого названия: он останавливает синтез пигмента на определенном этапе развития волоса. Ген-ингибитор носил самые разные генетические обозначения: Inhibitor (I), Bleacher или Bleaching (Bl), Silvering (Sv). Это последнее наиболее популярно в настоящее время.
Однако действием одного лишь гена-ингибитора нельзя было объяснить, почему одни из обладающим их кошек оказываются шиншиллами, а другие – серебристыми тэбби. Тем более действием гена-ингибитора невозможно было объяснить существование золотых типпированных окрасов. В начале 90-х годов американским генетиком J. Jerome было введено понятие гена-ингибитора эумеланина, чей доминантный (или полудоминантный) аллель избирательно подавлял синтез черного пигмента, оставляя основание волоса желтым. Этот ген был назван Eraser (Er). На основе взаимодействия двух ингибирующих синтез пигмента генов Дж. Джером была предложена бигенная теория наследования золотых и серебристых окрасов.
Взаимодействие аллелей серебристого и золотого окраса согласно бигенной теории
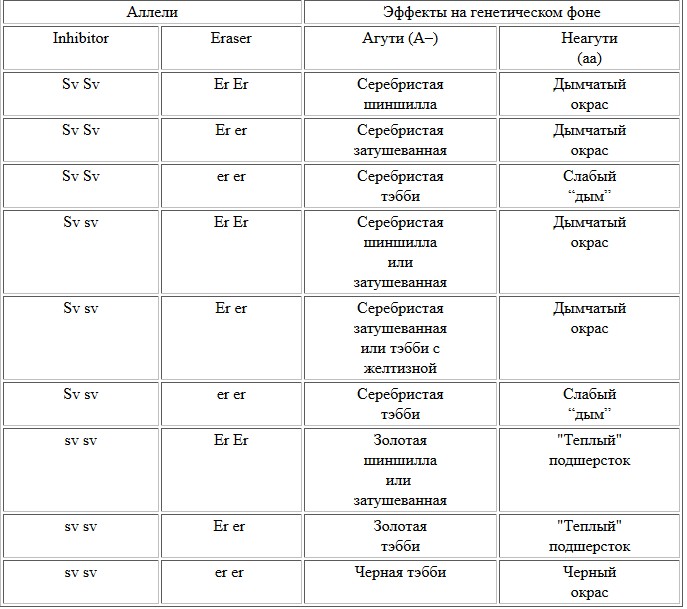
В середине 90-х . J. Wasselhuber было выдвинуто предположение, что феомеланиновое окрашивание прикорневой зоны у кошек золотого окраса возникает под действием не одного гена, а целого комплекса (полигена), названного ей “Widebanding” (Wb). Это название было дано по аналогии с геном Wide-band грызунов, формирующем широкую зону, лишенную полос, в основании волоса. “Желтая” полоса в основании волос (включая подшерсток) у кошек может наблюдаться как в сочетании с доминантным аллелем основного гена-ингибитора, так и без него. Для проявления типпированного окраса, – как золотого, так и серебристого, - наличие генов, определяющих “желтую полосу” (как бы они не назывались), оказывается совершенно необходимым. При формировании узкой бесполосной прикорневой зоны животные должны были выглядеть как тэбби.
Однако вопроса о генетическом разделении типпированных окрасов и тэбби бигенная теория – или теория о взаимодействии гена ингибитор с полигеном “широкой полосы” – окончательно не решала. Например, согласно этой теории золотые тэбби были гетерозиготами, а, следовательно, при спаривании между собой обязательно должны были давать не только золотых тэбби, но также шиншилл и затушеванных.
На роль генов, ответственных за такое разделение, были предложены аллели серии Тэбби. Тригенная теория в качестве необходимого для появления типпированных окрасов аллеля ввела аллель абиссинского окраса из серии Tabby - T a , который определяет отсутствие рисунка при тикированном окрасе волоса.
Но… последнее десятилетие фелинологического движения постепенно развернуло перед нами новую картину взаимоотношений между тикингом, типпингом и рисунком, причем картину весьма сложную. “За” участие в формировании типпированных окрасов аллеля, так или иначе связанного с исчезновением рисунка и появлением тикинга, говорит то, что среди потомства от скрещивания шиншилл (или затушеванных) с однотонными особями или с тэбби среди потомков часто появлялись животные тикированного окраса или же тэбби с одновременным присутствием рисунка и тикинга. Кроме того, среди кошек европейской короткошерстной или сибирской породы пятнистый или тигровый рисунки при золотистом окрасе почти всегда сопровождаются “избыточным” тикингом.
“Против” - то, что среди предков этих пород (персидская, британская, европейская, сибирская), естественно, никогда не было абиссинских кошек. Кроме того, форма проявления тикинга в этих породах никогда не достигает той степени выраженности, что характерна для абиссинских кошек - у животных, по крайней мере, в детском возрасте четко определяется рисунок, пусть и исчезающий с возрастом. Если признать таких особей гетерозиготными по доминантному (или полудоминантному) абиссинскому аллелю, то при скрещивании между собой они должны давать часть “абиссинских” потомков. За десять лет, в течение которых регулярно проводились подобные скрещивания, этого не наблюдалось.
С развитием пород восточного происхождения – ориентальной, сингапуры, цейлонской - ситуация с аллелем абиссинского окраса стала проблематичной. Оказалось, что тикированные тэбби существуют в двух вариантах – собственно абиссинском и “общем”. Этот последний, при котором корпус кошки не имеет рисунка, а ноги и хвост покрыты кольцеобразными полосами, характерен для таких пород, как ориентальная и сингапура. Иногда он встречается у аборигенных кошек Азии и Европы, особенно Восточной. У этих последних между “общим” тэбби и тэбби с рисунком можно найти все степени переходов, в том числе и пресловутый “избыточный” тикинг - без какого-либо наличия у них как абиссинских, так и типпированных предков. И все-таки окрас “тэбби без рисунка” традиционно рассматривался заводчиками как результат действия аллеля T a , наряду с другими аллелями того же локуса, определяющими разные формы рисунка.
А между тем накопившиеся данные об особенностях наследования “избыточного” тикинга, типпинга и “общего окраса тэбби” подводили некоторых фелинологов к предположению о том, что должен существовать некий ген (или группа генов) вне локуса тэбби, “убирающий” рисунок с корпуса животного. Причем данный ген должен был начинать свою работу уже после рождения котенка. Аналогию действия этого гена можно видеть у других представителей семейства кошачьих - льва и пумы, чьи котята рождаются пятнистыми.
Тэбби “без рисунка”
Проблемы наследования серебристого затушеванного и тикированного окрасов недавно были рассмотрены американской заводчицей, доктором ветеринарных наук Carol W. Johnson. Основой для ее теории, которую можно назвать "типпингом через тикинг", послужили серии скрещиваний, проведенные в собственном питомнике кошек американской короткошерстной породы (АКШ). Серебристый затушеванный окрас - один из старейших у АКШ, впервые короткошерстная “домашняя кошка” с таким окрасом зарегистрирована в CFA в 1913г. Чемпионский статус серебристые затушеванные АКШ получили в этой организации в 1951г. С 1993 г. признаны серебристые затушеванные голубые, черепаховые, голубо-кремовые, кремовые. А вот “тикированные тэбби” были первоначально признаны как “Другие вариации” (то есть без чемпионского статуса), затем, в 1993, были предложено их дисквалифицировать как “нехарактерный для породы окрас” – согласно утвердившемуся в фелинологии взгляду “все тикированные тэбби имеют абиссинское или восточное происхождение”. Между тем этот “нехарактерный окрас” столь регулярно выщеплялся среди АКШ, так что вопрос о генетике этих тикированных, или “бесполосных тэбби” приобрел для американских заводчиков принципиальное значение.
Поожим, для российских заводчиков в последние 5 лет он тоже становится весьма значимым. Среди потомства персидских, британских, сибирских кошек то и дело появляются тикированные тэбби "по восточному образцу" - то есть с полосатыми ногами и хвостом. Особенно часто такие звери появляются как результат работы по совершенствованию типа среди типпированных персов и британцев - при скрещиваниях между затушеванными кошками и особями окраса тэбби или затушеванных с однотонными особями. Эксперты не могут определить своего отношения к этим животным: с одной стороны, окрас счтается нетипичным для породы, с другой - понятно, что абиссинцев (или ориенталов) среди предков британцев или персов не было. В собственном питомнике К.Джонсон провела следующую серию скрещиваний. От серебристо-затушеванной кошки и красного мраморного кота были получены котята, имеющие рисунок на ногах, хвосте, шее и без рисунка или с крайне слабым (“теневым”) рисунком на теле (“почти затушеванными”). Следовательно, аллель, “убирающий” рисунок с корпуса животного, должен быть не полностью доминантным, а частично доминантным или ко-доминантным. Это верно и в отношении абиссинского аллеля T a. Но в потомстве от возвратного скрещивания мраморного кота с его “почти затушеванной” дочерью наблюдались три варианта тэбби – оба родительских: мраморные и “без рисунка”, но откуда-то появились еще и тигровые котята. Из этого распределения следовало, что в генотипах родителей присутствовали три аллеля - мраморного, тигрового и “бесполосого тэбби”. Но три этих аллеля не могут относиться к одному и тому же гену Tabby! Мраморный кот, естественно, в этом локусе несет два рецессивных аллеля мраморного окраса, а “затушеванная” черепаховая кошка-дочь должна быть носительницей всех 3 типов окраса тэбби (рецессивного мраморного, ко-доминантных “безрисуночного” и тигрового). Поскольку в одном локусе Tabby у одного животного может одновременно находиться только два аллеля, третий аллель должен относиться к другому локусу. Следовательно, аллель, который “стирает” рисунок и вызывает “общий тикированный” окрас, не входит в локус Tabby и не идентичен T a. К. Джонсон предложила назвать этот новый ген Unpatterned (с генетическим символом U). Этот аллель U определяет не частоту полос на волосе, но отсутствие рисунка на теле, то есть отсутствие различий в интенсивности окрашивания волос на различных участках корпуса. Действие этого аллеля маскирует проявление аллелей локуса Tabby. В гомозиготной форме он скрывает их действие почти полностью (за исключением слабых признаков рисунка на ногах и хвосте). У гетерозигот Uu обычно проявляется “теневой” рисунок на корпусе и четкий - на ногах и хвосте. Соответственно, при рецессивном генотипе животных - uu -будет проявляться тот тип рисунка, который заложен аллелями локуса тэбби.
Взаимодействие аллелей с учетом гена Unpatterned
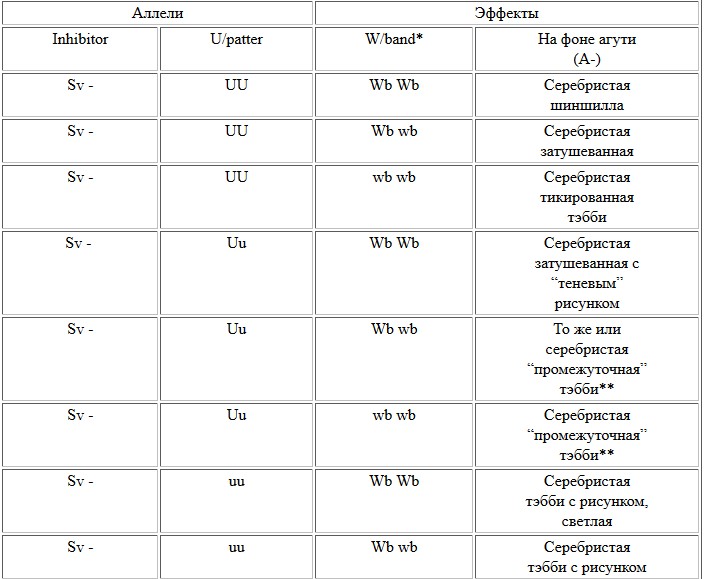
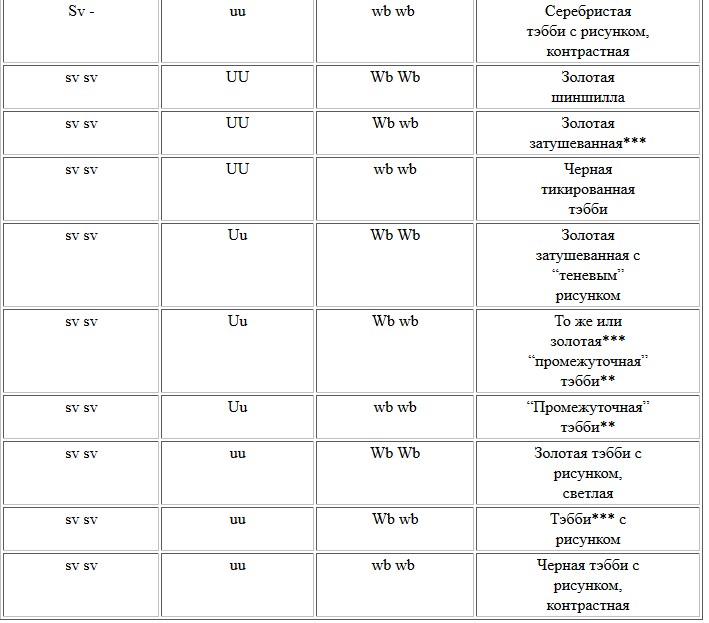
Widebanding здесь рассматривается как простой ген только условно. Реально, скорее всего, существует группа совместно действующих генов
** “промежуточный” тэбби – тэбби, у которого рисунок определяется, но недостаточно контрастен из-за значительного количества “избыточного” тикинга.
*** поскольку Widebanding все-таки полиген, гетерозиготные по генам Wb особи имеют разные варианты не полностью развитого “золота”, иногда с бурым или серым подшерстком.
Окончательно признать существование уникального гена “бесполосости” U пока что мешает наличие “переходных” между типпингом и тэбби окрасов. В поставленном нами скрещивании: мраморного кота и кошки золотого тикированного окраса с “остаточным” мраморным рисунком у потомков наблюдались обе родительские формы окраса и различные стадии перехода между ними – особи с достаточно четким рисунком, однако “засоренным” избыточным тикингом. Аналогичные картины наблюдаются и у БКШ при скрещиваниях серебристо-затушеванных особей с “рисунчатыми” тэбби – в потомства одновременно присутствуют и особи с крайне слабым “теневым” рисунком, и те, у которых рисунок определяется достаточно хорошо. Причем и у тех, и у других в большей или меньшей степени выражен одновременно “тикинг” и типпинг: нижняя часть волос рисунка обесцвечена, верхняя –покрыта черными и светло-желтыми полосами.
Итак, приходится признать, что решающим в “стирании” рисунка при формировании “бесполосых” тикированных тэбби и типпированных окрасов является (полу)доминантный ген U, отличный от генов Tabby. Данный ген довольно широко распространен в аборигенных популяциях кошек Восточной Европы, Сибири, Средней и Юго-Восточной Азии, и, вероятно, Северной Америки. Качественное развитие тикинга и типпинга, то есть исчезновение остатков рисунка зависит от минорных генов (модификаторов), действующих на фоне U.
Из такого предположения, видимо, и следует исходить господам фелинологам при определении своего отношения к регистрации "незаконных" тикированных окрасов в сложившихся породах.
Erase – гены “полосатых ног”?
Признак, попортивший немало крови любому заводчику шиншилл и затушеванных кошек, остаточные полосы на ногах и хвостах у типпированных животных. Гипотеза о существовании гена (или группы генов), “смывающей” остаточную полосатость на ногах животного, была впервые предложена Cathy Galfo в работе, посвященной затушеванным ориентальным кошкам. Для гена, удаляющего эти остаточные полосы, был предложен термин “Erase” (не путать с “Eraser”- одним из названий для гипотетического гена – ингибитора эумеланина у золотистых шиншилл!). Кажется, Erase наследуется независимо от гена U. Остаточный рисунок на ногах, хвосте и шее (“ожерелье”) может присутствовать при гомозиготности по аллелю U. Доказательством независимости гена(-ов) Erase от гена U можно считать окрасы типа “общий тэбби”, характерные, например, для сингапуры. В этой породе из поколения в поколение стабильно передаются “тэбби без рисунка” (на корпусе) в сочетании с полосатостью на ногах.
По нашим данным, среди британских короткошерстных кошек действительно соблюдается независимое наследование отсутствия рисунка на корпусе и наличия полос на ногах и хвосте. Степень выраженности последнего признака сильно варьирует (отмечались случаи “детской”, проходящей с возрастом полосатости). Вместе с тем наблюдалась значительная корреляция между общей “глубиной” типпинга, – иначе говоря, интенсивностью обесцвечивания волос (за счет активности основного гена-ингибитора меланина) и степенью выраженности полос. А вот корреляции между активностью ингибитора эумеланина (у золотистых шиншилл) и степенью развития рисунка на хвосте и ногах отмечено не было. В конечном счете, похоже, что рисунок на конечностях и хвосте действительно контролируется специфичной группой генов.
Chaos
Неравномерность типпинга и наличие размытых, теневых следов рисунка на корпусе животного Джонсон объясняет действием двух независимых друг от друга и гена U генетических факторов (или двух групп генов), названных ей Chaos и Confusion. Под Chaos Джонсон понимает формирование дополнительной темной полосы в светлоокрашенной (обесцвеченной) области волос рисунка. Этот эффект, проявляющийся у кошек с рисунком, называют у нас “избыточным тикингом”, в США - “тикингом классических (т.е. мраморных) тэбби”. Иначе выглядит проявление Chaos у затушеванных особей. Генетически “бесполосые” - uu - кошки, имеющие определенные аллели гена(-ов) Chaos, имеют тенденцию к проявлению “теневых” полос. И наоборот, другие аллели этого же гена(-ов) “убирают” эту остаточную полосатость с корпуса. “Теневая” полосатость отличается от “истинного” рисунка. Последний формируется за счет того, что волоски в рисунке имеют иное количество полос тикинга и иную картину их расположения, нежели волоски фона. Иногда же волоски истинного рисунка вовсе лишены полос тикинга. Волоски в “теневом рисунке” обычно имеют столько же полос, сколько и волоски фона (или и те и другие не имеют их вовсе), но протяженность темно-окрашенной зоны у волосков в рисунке изменена по сравнению с волосками фона. Генетически эти типы рисунка тоже отличаются - скрещивание двух особей с “теневым” рисунком может дать затушеванных котят, а двух тигровых особей - нет.
Confusion
У особей затушеванного окраса типпинг проявляется на всех волосках, но далеко не всегда оказывается однородным. Тщательное обследование показывает, что на одних волосках окрашен самый кончик, другие имеют 2 темных полосы, третьи прокрашены почти до половины. Встречаются даже чисто-белые и чисто-черные волоски. Изредка отмечается наличие тикированных волосков (число полос тикинга доходит до 7!). Часто присутствуют волосы смешанного типа, тикированные и типпированные одновременно - с белым основанием и тикированной верхней частью. Такое отсутствие координации между типпингом отдельных волосков было названо автором Confusion. Животные с высоким уровнем Confusion выглядят “загрязненными”. Небольшие степени Confusion обеспечивают различие в цвете затушеванных животных: наличие темной полосы вдоль хребта, светлых боков и белого живота. (см. блок).
Этот эффект никогда не наблюдается у новорожденных котят и развивается только с возрастом. У равномерно окрашенных (типпированных, затушеванных , тикированных) кошек или контрастных особях окраса тэбби производство пигмента в волосяных фолликулах тела или на отдельных его участках координировано во времени - они производят одно и то же количество пигмента, откладывающихся на одинаковых по длине участках волоса. По-видимому, ген(ы) Confusion нарушают эту координацию.
Действие Confusion не следует путать с так называемым Roaning - смешиванием окрашенных и белых волос. Такое смешивание порой наблюдалось в отдельных линиях у однотонных кошек и широко распространено у животных из других таксономических групп – собак и лошадей. Roaning наследуется как доминантный признак, сильно варьирующий по степени проявления, и совершенно независим от Confusion.
Confusion и Chaos у британских кошек
Верны ли наблюдения К. Джонсон для других пород кошек? Нами были обследовано несколько серебристо-затушеванных британских кошек как родственного, так и не родственного происхождения. Особи первой группы, происходящей от серебристо-затушеванного отца и серебристо-мраморной матери, в детстве имели легкий полосатый рисунок на корпусе. Впоследствии он проявлялся только как “теневой” - периодически проявлялся при разлиньке и исчезал при смене шерсти; в остальном же эти кошки выглядели как серебристо-затушеванные, с полосатыми ногами и хвостом. У таких особей максимальное разнообразие по цвету показывали покровные волоски на спине. Большая их часть была прокрашена на ?, причем около70% таких волосков имело три полосы тикинга (черный кончик - довольно узкая светло-желтая – широкая черная полосы). Именно эти покровные волосы создавали характерную для серебристо-затушеванного окраса темную полосу вдоль хребта. Меньшая часть покровных волос на спине была прокрашена на 1/3 – 1/8, без признаков тикинга. И, наконец, изредка попадались волоски, прокрашенные на 2/3 - 5/6 своей длины; эти волосы были тикированы (до 5 полос тикинга). Остепокровная шерсть на боках состояла в основном из волос, типпированных примерно на 1/6 – 1/8, без тикинга. Волосы с крайне узкой желтой полоской, прокрашенные на 1/3, попадались значительно реже, причем их распределение соответствовало “теневым” полосам рисунка. Наконец, остевая шерсть состояла только из типпированных на 1/8 волосков, а остепуховая была белой.
Вторая кошка, не родственная первым, признаков “теневого” рисунка не проявляла, как и полос на хвосте и ногах. Выглядела она равномерно типпированной и очень светлой. Покровные волосы на спине были типпированы на 1/6 – 1/8, причем последние преобладали. Покровный и остепокровный волос на боках был типпирован на 1/8, тикинга не наблюдалось.
Третья кошка, дочь предыдущей, также не родственная особям из первой группы, “теневой” рисунок проявляла только в детстве, во взрослом возрасте признаки рисунка отсутствовали полностью. Ее типпинг производил впечатление ровного, но “глубокого” - то есть шерсть выглядела относительно темной для серебристо-затушеванного окраса. Определялась более темная полоса вдоль хребта. Покровная шерсть на спине и на боках этой кошки оказалась типпированной на протяжении от 1/3 до 1/8, с преобладанием глубоко прокрашенных волос. Из этих последних многие имели 3 полосы тикинга, но средняя светло-желтая была крайне узкой. Остепокровная и остевая шерсть была типпирована на 1/6 – 1/8, без тикинга.
В целом распределение тикинга и типпинга у британских шиншилл подтверждает данные, полученные К.Джонсон на АКШ. Правда, пока что мы не нашли доказательств того, что Confusion и Chaos – независимо наследуемые группы генов, поскольку в нашем случае количество неравномерно прокрашенных волос коррелировало и с “теневым” рисунком и с неравномерностью типпинга - чего, кстати, нельзя было сказать о тикинге.
Иллюзия типпинга и стратегия разведения
Если наличие доминантного аллеля агути обеспечивает наличие зонарности волоса, то на распределение и количество этих зон (полос) влияют уже другие гены, которые К. Джонсон называет гены “частот зонарности” (banding frequency). “Частота зонарности” может быть определена как количество и величина чередующихся полос на волосе у животных окраса агути. Затушеванный окрас характеризуется единственной широкой эумеланиновой (черной) полосой на кончике волоса, в то время как абиссинские тэбби обладают множественными чередующимися эу- и феомеланиновыми полосами на всем протяжении волоска. Вероятно, гены “частот зонарности” регулируют временные интервалы отложения белков (пигментов). Одинаково выглядящие серебристо-затушеванными кошки на самом деле могут обладать различными “частотами зонарности”.
Признаки, контролируемые группами генов-модификаторов, хорошо поддаются целенаправленной селекции. Но стратегия племенного отбора животных в питомниках, направленная на улучшение качества серебристых типпированных окрасов, может быть строиться по-разному. Так, К.Джонсон выделяет три возможных варианта.
Вариант 1 - отбор против “частот зонарности” (то есть на уменьшение количества полос до тех пор, пока не остается единственная “концевая” черная полоса, то есть черный кончик волоса).
Вариант 2 - отбор на увеличение депигментации под действием непосредственно гена-ингибитора или генов “широкой полосы” в основании волоса, пока увеличенная депигментированная зона не закроет излишние полосы тикинга на волосе.
И вариант 3 - отбор на расширение всех полос, пока расширяющаяся вторая от кончика черная зона опять-таки не скроется под прикорневой депигментированной полосой.
Замечу от себя, что отбор этого типа должен быть сопряжен с достаточно высокой активностью генов-ингибиторов меланина и/или феомеланина. Варианты 2 и 3, наиболее частые в разведенческой практике, Джонсон характеризует как “иллюзию типпинга”, поскольку генетически волос остается зонарным, а не типпированным. Это утверждение выглядит замечательно верным. С последствиями различия стратегий отбора в разных питомниках мы сталкиваемся в любой породе: вспомните, сколько раз от пары типпированных животных рождались потомки со смешанным, тикированно-типпированным окрасом! Это – результат генетического несоответствия при одинаковом внешнем виде животных-родителей. Допустим, типпированный окрас кота вызван истинным отбором на снижение частот зонарности, а точно такой же на вид окрас кошки – отбором на увеличение депигментации. У потомка же частоты зонарности соответствуют материнским, а степень депигментации снижена под действием генов, пришедших от отца. Вот и “вылезает” на шкурке такого котенка тикинг пополам с типпингом, а заводчик в недоумении разводит руками…
“Сусальное золото”
Особенную остроту вопрос о стратегии отбора получает в разведении золотистых типпированных кошек. В отдельных питомниках, работающих с кошками этих окрасов, мы можем наблюдать четвертый вариант отбора – отбор на расширение одной из желтых полос тикинга (как правило, это первая сверху или же вторая снизу полоса). В присутствии генов Wb расширившаяся желтая полоса тикинга сливается с прикорневой феомеланиновой зоной, а кончик остается “типпированным”. Однако в результате скрещиваний таких животных с серебристыми партнерами потомки могут утратить ген Wb. Окрас подобных особей на первый взгляд весьма схож с обычным “золотом” – пока не смотришь на корни волос и подшерсток. Цвет этого последнего варьирует от серого до бурого, а основания волос – это касается в первю очередь покровной и остепокровной шерсти – тикированы (до 4-х полос тикинга). Центральную же часть волоса (около 2/3 –3/4 его длины) занимает расширенная желтая полоса, которая и придает кошачьей шерсти теплый “золотой” тон.
Формально говоря, с генетической точки зрения этот окрас золотистым не является. С точки зрения стандартов, его в лучшем случае можно определить как “плохой золотой”, так как стандартом оговорено, что серый и буорый подшерсток для золотых окрасов являются существенным недостатком (штраф вплоть до лишения титула). Но, даже если экспериты ужесточат требования к окрасу, такие “позолоченные” кошки неизбежно будут появляться в питомниках и на выставках – во всяком случае, до тех пор, пока заводчики при подборе пар не начнут учитывать, каким именно вариантом отбора на совершенствование типпинга создавались окрасы их племенных животных.
Заключение
Итак, если не все, то часть тикированных окрасов формируется не счет аллеля абиссинского тэбби, а за счет аллелей полудоминантного гена (или группы генов) U. Этот же ген играет существенную роль в развитии типпинга. Формирование типпированного окраса требует: присутствия: доминантного аллеля агути-фактора A; нескольких генов-ингибиторов меланина (полигенного комплекса ингибитора черного пигмента, а для серебристых окрасов еще и основного гена-ингибитора меланина Sv); (полу)доминантного аллеля гена “бесполосого тэбби” U; по крайней мере трех наборов полигенных комплексов, убирающих остаточный рисунок (“теневой” и полосы на ногах и хвосте), и полигенов, влияющих на число и ширину полос на волосе. Скрещивания мраморных тэбби с серебристыми затушеванными особями доказывают, что эти группы генов наследуются независимо друг от друга:
-----АА UU [WbWb][erase] [chaos] [confusion] [banding frequency] длязолотых
----- SvSv (или II, BlBl) АА UU [WbWb][erase] [chaos] [confusion] [banding frequency]
длясеребристых. типпированныхокрасов.
Инна Шустрова, кандидат биологических наук,
эксперт AB WCF, МКЭФ, RUI, гостевой МФА.